Researchers Implemented Reciprocal Conversion Between Annual and Polycarpic Perennial Flowering Behavior in the Brassicaceae
The cultivation of perennial crops offers significant potential for sustainable agriculture and addressing future climate change. In 2022, the development of perennial rice was highlighted as the world’s second top scientific breakthroughs by Science magazine. It is generally believed that perennial plants are more ancient, and annual plants have evolved from perennials. However, the genetic basis of the evolutionary transition from perennial to biennial and annual plants is still poorly understood.
On May 28, 2024, a research group led by Dr. WANG Jia-Wei at the CAS Center for Excellence in Molecular Plant Sciences had their research paper entitled “Reciprocal conversion between annual and polycarpic perennial flowering behavior in the Brassicaceae” published online in Cell.
This research strategy of this study is employing plant genera with rich variation in life-history strategies as models. Generation of cross-species mapping populations and forward genetics approaches were subsequently used to identify key genes determining the evolution of plant life-history strategies (i.e., polycarpic perennial flowering behavior). For this purpose, two pairs of compatible annual/perennial plant combinations were selected from the Brassicaceae family. Within the Crucihimalaya genus, Crucihimalaya himalaica is a polycarpic perennial, whereas Crucihimalaya wallichii exhibits facultative winter annual/polycarpic perennial characteristics. In the Erysimum genus, Erysimum nevadense is a polycarpic perennial. By contrast, Erysimum cheiranthoides is an annual. Using the two mapping populations with distinct life-history phenotypes from these four plants, three genetic regions associated with the evolution of life-history strategies were identified. According to the gene functional annotations of the model plant Arabidopsis thaliana, three closely related MADS-box transcription factor genes, namely FLOWERING LOCUS C (FLC), FLOWERING LOCUS M (FLM) and MADS AFFECTING FLOWERING (MAF), have been identified.
To further confirm the mapping results, FLC, FLM, and MAF loci in Crucihimalaya plants were mutated using CRISPR/Cas9 technology. By combining mutant alleles in different combinations through crossing, Dr. WANG Jia-Wei and his colleagues successfully recapitulated the evolutionary trajectory of plants transitioning from polycarpic perennial to biennial and annual: when FLC , FLM , MAF all remain functionally intact, the plant exhibits a robust polycarpic perennial phenotype; the absence of one or two of these genes leads to the adoption of facultative life-history strategies (i.e., weak polycarpic perennial, biennial, or winter-annual flowering behavior); when all these genes are completely inactive, the plant transitions to annual flowering growth habit.
Through RNA-seq and H3K27me3 ChIP-seq experiments on the parental and F1 generation plants, it was observed that FLC-like MADS-box genes in annual E. cheiranthoides tend to be stably repressed after vernalization, whereas homologous genes from perennial E. nevadense tend to be reset after vernalization. This difference constitutes the fundamental molecular basis underlying the establishment of polycarpic perennial life-history strategy and is determined by the specific sequences of the genes themselves. Notably, while FLC, FLM and MAF have close genetic relationships, they differ in terms of gene function, expression levels, as well as the timing and intensity of resetting.
Based on the experimental results mentioned above, Dr. WANG Jia-Wei and his colleagues propose how that the life-history strategies in Brassicaceae are evolved. Specifically, the transition among perennial, biennial, and annual flowering behavior is a continuum and determined by gene dosage effects of FLC-like MADS-box genes. The diversity of these genes in expression patterns, protein functions and epigenetic resetting modes, as well as their different combinations, allows plants to have a wide range of life-history strategies to adapt to diverse environmental conditions.
Finally, by introducing a perennial FLC gene into annual A. thaliana, Dr. WANG Jia-Wei and his colleagues successfully transformed A. thaliana from monocarpic annual to polycarpic perennial. This result suggests that A. thaliana has the prerequisite conditions of being polycarpic perennial. Perennial FLC-like MADS-box genes are likely necessary and sufficient for the establishment of polycarpic perennial life-history strategy. This finding thus lay the groundwork for future cultivation of polycarpic perennial Brassica napus.
Dr. ZHAI Dong, doctoral student ZHANG Lu-Yi, Dr. LI Ling-Zi and Dr XU Zhou-Geng from the CAS Center for Excellence in Molecular Plant Sciences are co-first authors of this paper. Dr. WANG Jia-Wei is the corresponding author. Dr. LIU Xiao-Li, Dr. ZHAO Bo, Dr. GAO Jian, Dr. WANG Fu-Xiang and doctoral student SHANG Guan-Dong also participated in this work. This research was supported by the New Cornerstone Science Laboratory, the National Natural Science Foundation of China, the Strategic Priority Research Program of the Chinese Academy of Sciences and the XPLORER PRIZE.
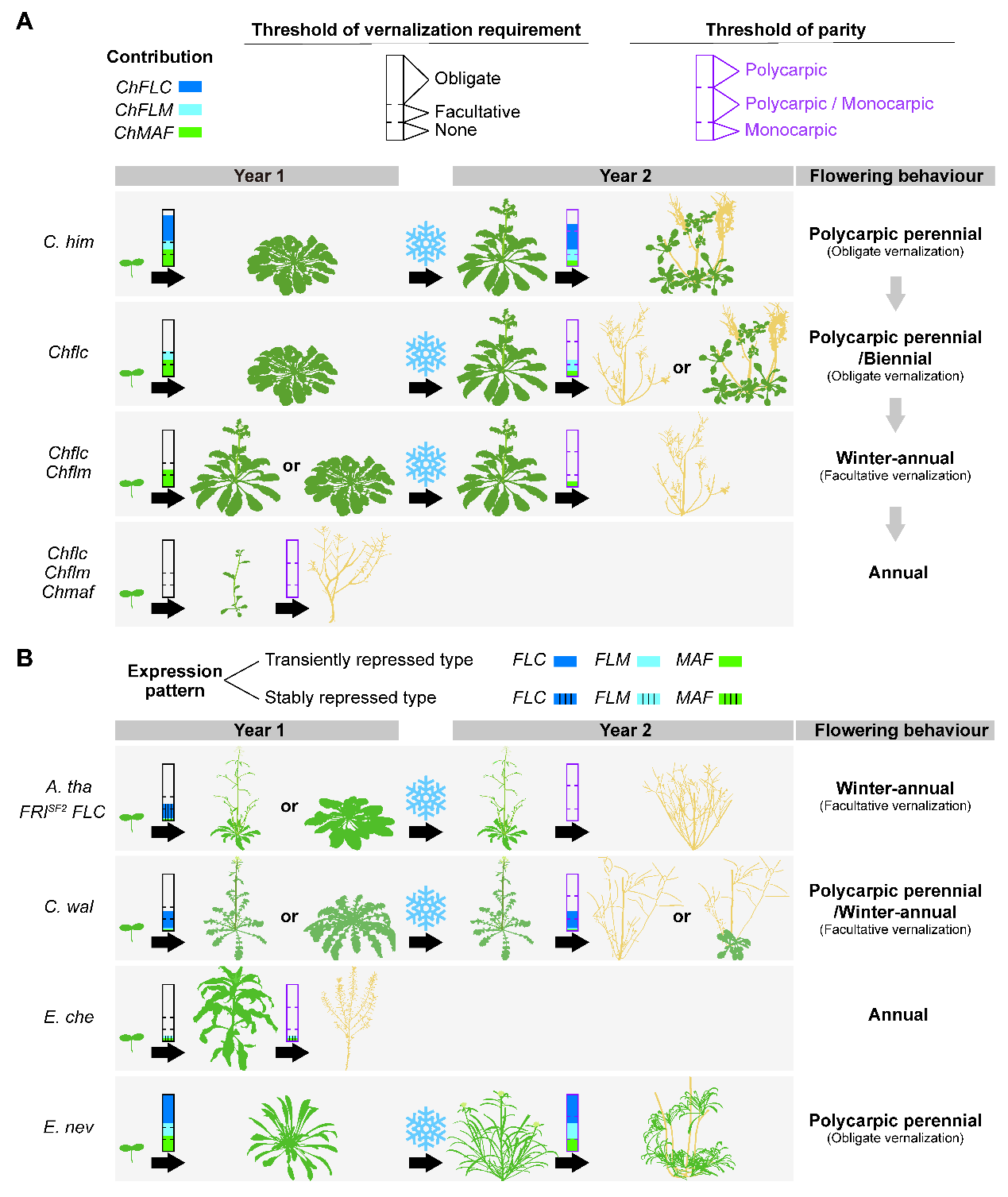
The dosage effect of FLC-like Genes determines the diversification of life-history strategies in Brassicaceae
(A) Regulation of life-history strategies in Crucihimalaya himalaica. Life-history strategies are primarily determined by two factors: vernalization requirement and parity. Vernalization can be categorized into obligate vernalization, facultative vernalization, and none. For parity, the categories include polycarpic, monocarpic, and facultative (polycarpic/monocarpic). In Crucihimalaya himalaica, FLC, FLM, and MAF are three homologous MADS-box transcription factors that inhibit flowering; their expression is suppressed by vernalization and reset after vernalization. Moreover, these three genes contribute differently to vernalization requirements and parity (indicated in the diagram by blue, cyan, and green, with no color representing no contribution). The combined effects of these genes determine the life-history strategies, the thresholds for vernalization requirements (black solid box) and parity (purple solid box) are depicted by dashed lines within the two solid boxes. In the diagram, C. him refers to Crucihimalaya himalaica, the wild type is polycarpic perennial. When the ChFLC gene is knocked out, the plants become perennial or biennial, with obligate vernalization requirement. Further knockout of the ChFLM gene turns the plants into winter-annuals with facultative vernalization requirement. When all three genes, ChFLC, ChFLM, and ChMAF, are absent, Crucihimalaya himalaica becomes an annual plant. Yellow in the diagram indicates whole or part of the plant tissues have withered and died. For convenience, the diagram only shows two consecutive years of life history.
(B) Regulation of life-history strategies in other Brassicaceae plants. Among these, A. tha FRISF2 FLC represents a winter annual Arabidopsis thaliana; C. wal refers to the weakly perennial plant (i.e., facultative perennial and winter-annual) Crucihimalaya wallichii; E. che refers to the annual plant Erysimum cheiranthoides; E. nev refers to the polycarpic perennial plant Erysimum nevadense. The expression patterns of FLC, FLM, and MAF differ among these plants, generally categorized into two types: stably repressed type (solid color with vertical lines) and transiently repressed type (solid color). The different expression patterns and contributions of these three homologous MADS-box genes determine the various life history strategies (refer to diagram A).
Contact:
Dr. WANG Jia-Wei, Professor
National Key Laboratory of Plant Molecular Genetics, Center for Excellence in Molecular Plant Sciences, Chinese Academic of Sciences





